外周血单核细胞活化抑制剂在制备治疗急性肝衰竭的药物中的用途的利记博彩app
本发明属于医学领域,具体涉及外周血单核细胞活化抑制剂在制备治疗急性肝衰竭的药物中的用途。
背景技术:
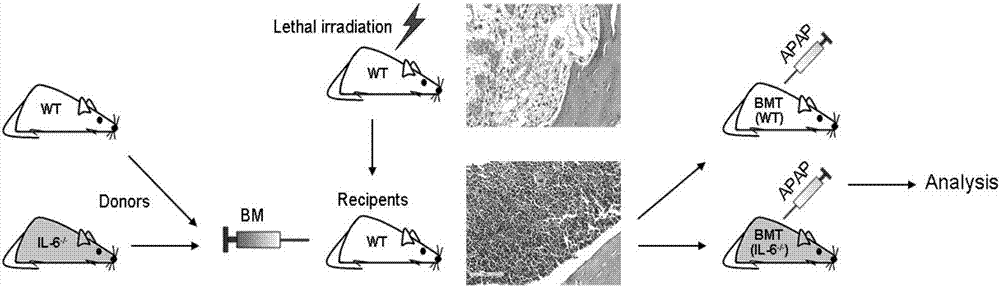
:急性肝衰竭(acuteliverfailure,alf)发病急骤,死亡率高,危害极大。在发达国家,alf发病率不到百万分之十,但在包括中国在内的发展中国家,尽管缺乏准确数据,但alf发病率显然高出许多。2013年bernalw等在《新英格兰医学杂志》发表综述,沿用了1970年提出的alf定义:无基础肝脏疾病的严重的、具有潜在可逆性的肝脏损伤,在出现首发症状后8周内发生肝性脑病。根据首发症状与肝性脑病之间的间隔,又可分为超急性、急性、亚急性,等等。之后世界各大肝病研究机构相继发布了多个alf诊疗指南,并对alf的定义进行相应的修订。我国肝病学家在2012年《肝衰竭诊治指南》中提出将2周内出现以ⅱ期以上肝性脑病为特征者定义为alf。我国alf病因与西方国家差异显著。国外的alf定义强调本次发病为“孤立”的,突发的,此前并没有肝脏基础疾病,以药物性肝损伤为主。而我国alf的常见原因仍然是病毒性肝炎或合并有肝炎。事实上,随着人群中hbv携带率的下降,我国alf病因也在发生着深刻变化。解放军302医院肝衰竭诊疗与研究中心从7家三级医院纳入177例alf患者进行研究,在病因中,药物毒性占43.5%,仅11.3%的alf由急性病毒性肝炎所致(大部分为hbv感染,其余包括hev、hav、cmv和ebv感染等)。据该报告,我国alf的主要原因已不再是急性病毒性肝炎,而是与发达国家类似,以药物性肝损伤为主。急诊肝移植是alf有效治疗手段,常规内科治疗死亡率极高。在上述研究中,所有纳入病例均未接受肝移植,死亡率高达63.82%。受限于供肝的极度短缺,我国每年肝移植适应症患者如愿获得供肝的比例不到1%。alf治疗策略主要包括去除病因、促进肝再生、肝功能支持、抗炎症反应以及对症支持等,但效果均不甚满意。组织工程肝、生物人工肝和肝细胞移植可能对alf有效,但技术尚欠成熟、缺乏足够临床病例积累和循证医学证据,尤其是缺乏大样本随机临床对照试验,仍然难以作为常规治疗手段,alf的治疗依然是世界性的难题。技术实现要素:为了解决上述问题,本发明提供了一种新的治疗和/或预防急性肝衰竭的药物。外周血单核细胞活化抑制剂在制备治疗和/或预防急性肝衰竭的药物中的用途。其中,所述外周血单核细胞活化抑制剂是抑制外周血单核细胞的il-6表达水平和/或ccr2表达水平的物质。外周血单核细胞:是血液中一群体积最大的白细胞,来源于骨髓,是脊椎动物先天性免疫系统重要组成部分,也参与适应性免疫功能,能分化为成熟的巨噬细胞或髓系树突状细胞。优选地,所述抑制剂是脐带间充质干细胞。优选地,所述抑制剂是il-6基因敲除型单核细胞。优选地,所述抑制剂是il-6基因沉默剂。本发明还提供了一种治疗和/或预防急性肝衰竭的药物,它是以外周血单核细胞活化抑制剂为活性物质,加上药学上可接受的辅料或者辅助性成分制备而成的制剂。其中,所述外周血单核细胞活化抑制剂是抑制外周血单核细胞的il-6表达水平和/或ccr2表达水平的物质。其中,所述抑制剂是脐带间充质干细胞、il-6基因敲除型单核细胞或者il-6基因沉默剂。其中,所述制剂为注射制剂。本发明还提供了药物在制备治疗治疗和/或预防急性肝衰竭的药物中的用途。本发明实验证实了抑制单核细胞的活化能够有效治疗和预防急性肝损伤,脐带间充质干细胞验证了抑制单核细胞的活化的确能够有效治疗和预防急性肝损伤,说明单核细胞活化抑制剂可以有效治疗和预防急性肝损伤,可以作为治疗和预防急性肝损伤的药物使用,其中,单核细胞活化抑制剂可以是il-6基因敲除型单核细胞,也可以是脐带间充质干细胞,还可以是其他抑制单核细胞活化的物质,如il-6基因沉默剂。本发明为临床治疗急性肝损伤提供了一种新的选择,应用前景优良。显然,根据本发明的上述内容,按照本领域的普通技术知识和惯用手段,在不脱离本发明上述基本技术思想前提下,还可以做出其它多种形式的修改、替换或变更。以下通过实施例形式的具体实施方式,对本发明的上述内容再作进一步的详细说明。但不应将此理解为本发明上述主题的范围仅限于以下的实例。凡基于本发明上述内容所实现的技术均属于本发明的范围。附图说明图1小鼠尾组织pcr基因型鉴定结果图2小鼠骨髓移植实验流程图。图3wt小鼠x射线全身辐照后骨组织清髓he染色(左为正常骨髓,右为清髓后)。×400图4股骨组织he染色鉴定骨髓重建。左:wt小鼠来源全骨髓移植重建1月后股骨he染色;右:il-6-/-小鼠来源全骨髓移植重建1月后股骨he染色。×400图5apap诱导肝损伤后肝脏he染色,显示bmt(il-6-/-)组小鼠肝损伤更为轻微,其修复也更迅速。×400图6apap诱导肝损伤后肝功生化指标检测。*p<0.05图7luminex液相芯片检测两组小鼠肝损伤后血清炎性细胞因子表达水平。*p<0.05图8fcm检测两组小鼠外周血单核细胞cd11b/ccr2的表达。***p<0.001图9外周血细胞因子的表达变化。*p<0.05,&p<0.01,$p<0.001图10毒素给予后一天动物外周血scd163表达水平。***p<0.001图11huc-mscs抑制c-mos向肝脏迁移。(a)毒素诱导猴alf后各时间点肝脏内kcs(cd68+)和招募的c-mos(mac387+)。scalebars=50μm。(b-c)定量统计肝小叶周围cd68+和mac387+两种巨噬细胞在各时间点的阳性数目,计数方法为每10个不同400倍视野(样品至少来自3只猴)进行计数并取其平均值。*p<0.05,**p<0.01,***p<0.001。图12免疫荧光双染检测肝内kcs增殖情况(图中未见增殖的kcs,而ki67阳性的细胞为增殖的肝细胞)图13fcm检测动物外周血cd14+cd16+ccr2+单核细胞。***p<0.001图14外周血cd14+cd16+ccr2单核细胞il-6mrna水平。***p<0.001图15alf猴第3天时肝脏cd45+炎症细胞浸润情况。图16预防性干细胞输入组实验流程图。图17预防性给予huc-mscs显著抑制肝细胞损伤及巨噬细胞活化。***p<0.001图18健康组与预防组动物外周血清scd163水平。图19fcm分析外周血cd14+cd16+ccr2+细胞和定量pcr检测il-6的mrna表达。图20fcm检测预防组白细胞群和treg细胞亚群数量的变化。图21预防组第2天和第5天外周循环中细胞因子水平。**p<0.01,***p<0.001具体实施方式主要缩写词表:实施例1抑制外周血单核细胞活化与急性肝衰竭的关系一、基因敲除的单核细胞显著改善急性肝损伤(一)材料1实验动物c57bl/6jil-6-/-kopf(il-6-/-)小鼠由四川大学华西医院余希杰教授惠赠。小鼠分笼饲养于spf级动物房,每天12h光照,自由饮水和进食。由四川大学华西医院实验动物中心(持一级动物饲养合格证)按照“四川省医学类实验动物饲养和管理条例”及“实验动物管理条例”进行喂养和处理。敲除il-6基因可以抑制单核细胞的活化,并且骨髓细胞中,只有单核细胞会表达il-6,通过骨髓移植il-6-/-敲除小鼠的骨髓的方式可以有效考察单核细胞活化与疾病之间的关系。2主要实验试剂1)小鼠基因型鉴定相关试剂:pcr试剂、琼脂糖、电泳缓冲液、核酸染料等2)全骨髓获取和移植相关试剂:75%乙醇,rpmi-1640培养基,红细胞裂解液,3%台盼蓝等3)apap:n-乙酰对氨基酚(sigma,美国)4)组织固定、包埋及病理学染色相关试剂:甲醛、乙醇、二甲苯、石蜡、苏木素染液、伊红染液、浓盐酸(科隆试剂,成都)、油红o染料5)cd11b,ccr2流式抗体(bd,美国)3主要仪器1)x射线辐照仪x-rad320:(pxi,美国)2)骨髓分选相关手术器械3)电泳系统:bio-rad4)pcr仪:(ptc-200peltierthermalcycler,mjresearch)5)凝胶成像系统(mqx200,biotek,美国)6)luminex200(millipore,美国)7)流式细胞仪fc500(二)方法1基因型鉴定il-6基因敲除小鼠由遗传背景为c57bl/6的野生型小鼠将新霉素抗性基因插入il-6基因的第二外显子诱导il-6基因突变产生,得到特异性il-6-/-敲除小鼠(详细制备方法如lingxiangkong,deletionofinterleukin-6inmonocytes/macrophagessuppressestheinitiationofhepatocellularcarcinomainmice,kongetal.journalofexperimental&clinicalcancerresearch(2016)35:131doi10.1186/s13046-016-0412-1所示)。通过pcr检测鼠尾样品,确定小鼠的基因型。il-6小鼠基因型鉴定方法:1)剪取出生2-4周龄小鼠尾巴于ep管中(1-2cm)。2)加入75μlbuffera碱溶液,100℃沸水煮40-45min。3)加入225μlbufferb酸溶液中和buffera,得到小鼠基因组dna模板溶液。4)按照表1配置反应体系,总体积20μl。引物序列如表2所示。基因分型buffera(ph≈12.0):100μl的0.5medta(ph8.0)、625μl的10mol/l双蒸水定容至250ml。基因分型bufferb(ph=3.0):浓度为40mm的trisbase,浓盐酸调ph值至3.0。表1.基因分型pcr反应体系(20μl)根据以下条件进行pcr反应:表2.基因分型pcr引物配置3%琼脂糖凝胶,配制100mltae:琼脂糖粉(agrose)2g,加入三角烧瓶的tae溶液混匀,微波炉加热煮沸至完全溶解;加入8μlgoldviewdna染料轻轻混匀,溶液室温冷却后倒入电泳槽中,放入梳子,待胶冷却凝固;电泳时,加8μl反转录产物样品,130v电压下电泳25min;电泳结束后,取出,凝胶成像系统紫外光下观察及采集图像。2骨髓移植(bonemarrowtransplantation,bmt)1)骨髓移植受体为c57bl/6野生型(wt),小鼠x射线辐照清髓处理:spf级,8-12周成年鼠;致死剂量辐照清髓:小鼠在辐照清除骨髓前一周,即开始饮用添加抗生素(庆大霉素320000u/l和红霉素250mg/l)的酸化水,全身辐照总剂量为8.0gy,剂量率为0.5gy/min;2)全骨髓细胞制备:il-6-/-或wt小鼠颈椎脱位处死后,置于75%的乙醇中浸泡10min,超净台无菌条件下剥离股骨和胫骨;将所取骨头转移至预先加入rpmi-1640培养基的研钵内,冰上轻轻研磨数分钟后,200μm滤器过滤,离心1000rpm/10min;去上清,加入红细胞裂解液,裂解10min,离心1500rpm/10min;去上清,加入1mlrpmi-1640重悬细胞,洗涤,台盼蓝鉴定活性不小于95%,计数,调整骨髓细胞浓度2×107/ml(该骨髓细胞中的单核细胞为il-6基因敲除型单核细胞),备用;3)骨髓移植(bmt):将分离所得的il-6-/-或wt全骨髓细胞以2×107/只,经尾静脉移植入受体wt小鼠体内。3apap诱导建立小鼠急性肝损伤(ali)模型骨髓移植存活一个月后的小鼠用来诱导肝损伤模型。小鼠分组情况:野生型骨髓重建组和il-6-/-基因缺陷小鼠骨髓重建组,每组分别6只。所有小鼠均禁食不禁饮水12h后,腹腔注射300mg/kg的apap溶液,注射apap后恢复自由进食和饮水。4he染色预计时间点获取小鼠肝脏行he常规染色,染色步骤同前。骨组织he染色中涉及的脱钙方法和步骤:(1)10%中性福尔马林固定骨股组织4-5天,流水冲洗标本20min;(2)使用10%edta脱钙液进行脱钙处理后常规he染色。5小鼠肝功能检测采用乙醚吸入麻醉小鼠后,眼球取血至ep管中,离心1200rpm/10min,取上清送检肝功检测,包括:alt,ast和tbil。6小鼠血清细胞因子检测方法简述如下:提前20min从冰箱中取出试剂盒及冻存的样本,以平衡室温。在板上的每个孔中加入200ul的洗涤液。封板,室温(20-25℃),放在振荡器上10min清洗。倒掉洗涤液,把板反过来在可吸附的纸上轻叩几次以去掉残余物质。在适当的孔中加每种标准品和质控品,用分析缓冲液作为(本底)。在背景孔和所有标本孔中加入25ul分析缓冲液。在本底、标准品和质控品中加入25ul基体溶液。加入25ul标本到适当孔中。涡旋混合瓶,每个孔中加入25ul混合的珠子,在加珠子过程中间歇的摇动装珠子的瓶子,避免沉淀。用封板膜封板,在板的外面上铝箔,放在振荡器上室温(20-25摄氏度)振荡孵育2h或者4℃孵育过夜。轻轻地去掉板里的内容物,洗板两次。在每个孔中加入25ul检测抗体。封板,包上铝箔,在振荡器上室温振荡孵育1h。在每个含有25ul检测抗体的孔中加入25ul链霉亲和素-藻红蛋白。封板,包上铝箔,在振荡器上室温振荡孵育30min。轻轻地去掉板里的内容物,洗板两次。在所有孔中加入150ul鞘液。在振荡器上重悬珠子15min。将板放在flexmap上用xponent运行。保存并使用样条曲线拟合法分析中间荧光强度数据,计算标本中细胞因子的浓度。7小鼠外周血单核细胞ccr2表达小鼠眼球取血(体积约为200μl)至肝素钠洗涤过的ep管,与肝素钠混匀后,直接加入cd11b,ccr2流式检测抗体,室温孵育20min,流式细胞仪检测。(三)结果1小鼠基因型鉴定对特定基因片段扩增基因型鉴定结果(图1)显示,il-6基因缺陷(il-6-/-)小鼠pcr产物扩增片段为1400bp,野生型(wt)扩增片段为900bp。2小鼠骨髓移植实验流程骨髓移植实验流程如图2:wt野生型小鼠辐照清髓6h内,使用wt和il-6-/-不同基因型来源小鼠骨髓重建清髓小鼠骨髓系统。骨髓重建成功后,再经apap诱导建立急性肝损伤模型。3wt和il-6-/-来源的全骨髓均可重建wt小鼠骨髓造血系统wt野生型小鼠股骨组织清髓he结果显示,经x射线辐照清髓后,骨髓腔中骨小梁附近的的骨髓细胞成分几乎完全被摧毁(图3)。he染色结果显示wt或il-6-/-基因型来源的全骨髓移植1月后,均可重建辐照清髓小鼠的骨髓造血系统(图4)。4wt和il-6-/-来源骨髓重建后的小鼠apap诱导肝损伤情况apap腹腔注射1天后,he染色结果显示两组动物肝脏中央静脉区域出现不同程度的坏死区域(图5)。其中,wt小鼠来源骨髓受体小鼠,记做bmt(wt)小鼠,其坏死区域面积显著大于il-6-/-小鼠来源骨髓受体小鼠,记做bmt(il-6-/-)小鼠,提示其肝损伤更为严重。第2天时bmt(wt)小鼠肝脏仍表现为大片状的坏死区域,而bmt(il-6-/-)组的坏死区域逐渐修复消退,仅表现出嗜酸性和水肿变性。表明在apap同等剂量诱导的急性肝损伤条件下,bmt(il-6-/-)组小鼠的肝损伤情况显著轻于bmt(wt)组。肝功能生化指标也验证了上述组织病理学结果。apap诱导第1天和/或第二天,bmt(wt)组血清中转氨酶alt、ast和tbil水平显著高于bmt(il-6-/-)组。之后两组肝功生化指标均回落,第3天时都逐渐恢复至接近正常水平值(图6)。以上结果表明,接受il-6基因剔除的骨髓重建小鼠,apap诱导急性肝损伤条件下表现出较接受wt骨髓组更轻的肝损伤表现。5小鼠血清炎性细胞因子检测bmt(wt)和bmt(il-6-/-)两组小鼠血清中炎性细胞因子的表达水平结果如图7所示。bmt(il-6-/-)组il-6表达水平在各时间点均显著低于bmt(wt)组。apap诱导肝损伤1天和2天后,bmt(il-6-/-)组炎性细胞因子tnf-α、ifnγ和趋化因子mcp-1表达水平显著低于在bmt(wt)组,两组有显著性差异,p<0.05;第3天时随着肝损伤的修复,炎性因子表达水平回落。6流式细胞术检测小鼠外周血单核细胞ccr2的表达接下来我们分析了两组不同基因型骨髓来源的小鼠在肝损伤时外周单核细胞(c-mos)ccr2的表达水平。如图8所示,bmt(il-6-/-)组小鼠c-mosccr2的表达的确被显著抑制,低于bmt(wt)组2倍多,具有显著性差异,说明敲除单核细胞的il-6基因,可以有效抑制单核细胞的活化。(四)讨论我们使用il-6基因缺陷小鼠来源的骨髓重建辐照清髓的野生型小鼠,观察il-6基因缺陷的单核细胞在apap诱导肝损伤后,小鼠单核细胞活化情况。首先,il-6基因缺陷小鼠来源的骨髓和野生型小鼠骨髓均能够成功重建辐照小鼠的骨髓造血系统;其次,相同肝损伤诱导条件下,il-6基因缺陷骨髓重建的小鼠肝组织病理及肝功生化表现出较野生型更轻微的损伤。同时,炎性细胞因子表达水平亦显著低于野生型对照组。最后流式细胞术也发现外周血单核细胞的ccr2(ccr2为单核细胞活化的标志物)表达百分比显著低于对照组。以上结果表明,正是由于单核细胞il-6基因的缺失,导致了单核细胞活化受到抑制,进而导致了两种不同基因型来源骨髓重建小鼠肝损伤后不同的病理损伤表现。(五)结论il-6基因剔除小鼠来源的骨髓重建的小鼠与wt骨髓来源小鼠相比,由于其可以抑制外周血单核细胞活化,降低单核细胞il-6的表达水平,因而在同等剂量apap肝毒性药物诱导的急性肝损伤中,表现出较轻的肝组织病理损伤、炎性细胞因子水平以及更低水平的单核细胞活化状态。实验结果说明,通过抑制外周血单核细胞的活化,可以有效治疗和预防急性肝损伤,具体地,可以使用il-6基因敲除型单核细胞替代外周血中的正常单核细胞,从而达到治疗和预防急性肝损伤的目的。二、间充质干细胞通过抑制单核细胞活化来改善急性肝损伤(一)材料1恒河猴及人脐带间充质干细胞。2恒河猴外周血清、肝穿刺活检组织及猴外周血单核细胞。3主要实验试剂1)免疫组化相关试剂:甲醛、乙醇、二甲苯、石蜡、苏木素染液、伊红染液、浓盐酸(科隆试剂,成都)等2)抗体:详见下表抗体及相关试剂信息3)luminex液相芯片试剂盒:acytokinemonkeymagnetic29-plexpanel(lifetechnologies)4)humancd163quantikineelisakit(r&dsystems,美国)5)ficoll淋巴细胞分离液(ge,美国)6)trizol(invitrogen,美国)7)pcr相关试剂:10×taqbuffer,taqdnapolymerase,25mmmgcl2dntp混合物(tiangen,中国)4琼脂糖凝胶电泳相关试剂琼脂糖,50×tae电泳缓冲液,核酸染料(goldview,赛百胜,北京)5主要仪器设备1)流式细胞仪fc500cytomics(beckmancoulter,fullerton,calif.,美国)2)巴德mg15-22全自动活检枪(bard,美国)3)超纯净水产生系统(milli-q,美国millipore公司)4)各型号移液器(eppedorf公司,德国)5)全自动脱水机(dako,美国)6)石蜡包埋机(bm2135,leica,德国)7)石蜡切片机(eq1150c,leica,德国)8)光学显微镜(020-518.500,olympus,日本),荧光显微镜(leica,德国)9)图像采集系统(olympusdpcontroller70,日本)10)微型低温离心机(thermolfisher,美国)11)pcr仪(ptc-200peltierthermalcycler,mjresearch)12)图像编辑软件(photoshopcs2,graphpad5.0,美国)13)统计分析软件(spss19.0,美国)14)流式细胞术分析软件(kaluzav1.20software,德国;cxp2.1流式软件,德国)15)超低温冰箱(thermoscientific公司,美国),液氮生物容器(成都液氮容器厂)全自动脱水机(dako,美国)16)luminex200system(millipore,美国)(二)方法1免疫组织化学(ihc)染色肝脏穿刺组织石蜡包埋完成后,5μm连续切片,捞片后将切片置于65℃烤箱中烤片6过夜。取出过夜烤好的切片,按照常规步骤进行脱蜡水化:二甲苯溶液10min×2,无水乙醇3min×2,95%酒精,85%酒精,75%酒精各3min,流水冲洗5min,蒸馏水洗3min×3次。ihc时,先用3%h2o2灭活内源性过氧化物酶,蒸馏水漂洗两次后用tris-edta(ph=9.0)抗原修复液进行抗原修复;然后用组化标记笔将待切片组织外围圈好,滴加事先已稀释好的一抗抗体于组织上,37℃孵育2h。pbs漂洗3次后滴加envision二抗,37℃孵育45min。pbs漂洗3次后使用dab显色液显色;苏木素染色液复染;脱水透明之后用中性树胶封片。2免疫荧光(if)染色1)将肝脏石蜡切片脱蜡水化(见上述步骤);pbs冲洗2次,每次5min;2)抗原修复:将切片湿盒浸泡在抗原修复液(柠檬酸盐缓冲液)中,微波炉30min,冷却到室温,pbs冲洗3次,每次5min;3)封闭:5%正常山羊血清工作液封闭,室温孵育15min,倾倒封闭液;滴加配制好的抗体工作液(cd68与ki67混合配制),4℃过夜;4)次日,复温后,pbs冲洗三次;滴加适量荧光标记二抗工作液(混合配制),室温孵育1h;pbs清洗三次,每次5min;5)dapi工作液染核5min后,清洗2次;6)缓冲甘油封片,荧光显微镜观察,采集图像。3elisa检测血清scd163使用人可溶性cd163试剂盒,此方法检测灵敏度范围为1.56-100ng/ml,为确保样本能够准确测出,将血清样本稀释100倍,之后按照试剂盒说明书指导下进行定量检测,具体方法:1)包被:每个聚苯乙烯反应孔中加入100μl稀释成工作液的兔抗cd163,4℃过夜,次日弃去孔内溶液,缓冲液洗涤3次,每次3min(简称洗涤,下同);2)加样:弃去孔内液体,洗涤。加入100μl用pbs稀释100倍的样品于反应孔中,同时做空白孔,阴性对照孔及阳性对照孔,室温孵育1h;3)加入酶标抗体:洗涤反应孔,加入100μl稀释的cd163单克隆抗体(1:500),室温孵育1h,洗涤,加入100μl过氧化酶标记的抗体(山羊抗小鼠免疫球蛋白,dakop447,1:4000),室温孵育1h,洗涤;4)加底物显色:先加入100μl过氧化氢/邻苯二胺盐酸盐底物溶液,再加入50μl的硫酸,在elisa酶标仪上于492/620nm波长处读取数值。批间精密度变异系数cv分别为6.3和9.6%,对应数值分别为1.84和3.91mg/l(n=19×2)。4恒河猴外周血单核细胞(monocyte)分离和纯化1)后腿隐静脉无菌采集约5ml外周血于肝素锂抗凝管中,室温2000r/min离心20min,移液枪吸弃上层液体,沉淀层为细胞。2)细胞沉淀层中按1:1体积比加入pbs缓冲液,混匀;取一新15ml离心管加入一定体积的ficoll淋巴细胞分离液;3)使用移液枪吸取混匀的细胞缓慢小心加入ficoll淋巴细胞分离液的上层,离心,结束后,可见离心管中液体分为4层。4)其中第二层云雾状,即主要为个单核细胞。吸弃第一层的血浆,收集血浆和第三层ficoll分离液之间的白膜层于新离心管中。5)加入5倍以上体积pbs吹打混匀,离心洗去所收集的单核细胞中的ficoll淋巴细胞分离液,共洗涤2次。6)rpmi-1640培养基以上述同样的离心条件洗涤1次,离心结束后,用小量程移液枪尽量吸走全部的上清。再用一定体积添加10%胎牛血清的rpmi1640重悬细胞。7)吸取50μl细胞悬液,血小板计数统计并调整细胞浓度至106/ml;浓度为2%台盼蓝染色检测细胞的活力;取一定体积的细胞悬液,测试单核细胞表面标志物cd14,以确保所得单核细胞的纯度。5流式细胞术检测单核细胞活化收集恒河猴外周血2ml于抗凝管中,离心,1500rpm,5min,弃掉上次血清;按照一定比例加入红细胞裂解液,8-11min左右;离心弃上清液,清洗下层细胞团。加入用pbs稀释的荧光素标记的anti-cd14、cd16、ccr2抗体200μl,移液枪轻轻吹打混匀,室温避光孵育15-20min;离心弃上清液,加入pbs继续洗涤2次;加入500μl冷pbs,移液枪混匀细胞,转移置流式管中,4℃冰箱保存,待测。6单核细胞总rna提取及il-6mrna水平检测提取mrna使用的移液枪枪头、ep管等用含0.1%焦炭酸二乙酯(depc)ddh2o浸泡过夜后,高温高压灭菌除去depc残留,100℃烘箱烘干备用。1)trizol裂解得到的单核细胞,提取rna:在离心管中加入1mltrizol溶液,移液枪吹打离心管底部的细胞团,充分裂解,室温下静置10min。2)离心管中加入氯仿:加入1/3trizol体积的氯仿,盖紧离心管,涡旋仪剧烈振荡30s,室温下静置5-10min,12,000×g,4℃离心15min。3)离心机中小心取出离心管,缓慢吸取上清液转移至新的离心管管中,注意不要吸出中间白色蛋白质层。4)向上清中加入等体积的异丙醇,轻柔上下摇晃数次至混匀,在室温下静置10-20min。12,000×g,4℃离心10min,离心管底部出现沉淀,即rna。5)轻柔弃去上清液,沿离心管管壁加入1ml75%的乙醇,轻轻上下颠倒离心管,洗涤管壁。12,000×g,4℃离心5min。轻柔倾倒乙醇,轻放室温干燥2-5min。6)加入适量的rnasefreewater溶解rna样品,待rna沉淀溶解后,立即进行反转录,剩余样本分装并冻存于-80℃冰箱。逆转录反应测定总rna的浓度和od260/od280值。按下表所示配制逆转录反应液(20μl体系),反应液配制全部在冰上进行。(3)根据以下逆转录程序进行rt-pcr:5minat25℃30minat42℃5minat85℃10minat10℃,反应结束后cdna样品放入-80℃冰箱长期冻存。逆转录pcr反应体系(20μl)荧光定量pcr按照下表配制qpcr反应液(20μl体系),所有操作反应全部在冰上进行。荧光定量pcr反应体系(20μl)根据以下条件进行qpcr反应:qrt-pcr引物购自上海生工生物技术有限公司,具体信息如下表所示。geneforwarprimersequencereverseprimersequenceil-65'atgaggacacttgcctggtg3'5'gctggcatttgtggttggtt'3'gapdh5'tcgagagtcagccgcattttc3'ggaacttgccatgggtggaa3'7luminex检测恒河猴血清细胞因子方法简述如下:提前20min从冰箱中取出试剂盒及冻存的样本,以平衡室温。在板上的每个孔中加入200ul的洗涤液。封板,室温(20-25℃),放在振荡器上10min清洗。倒掉洗涤液,把板反过来在可吸附的纸上轻叩几次以去掉残余物质。在适当的孔中加每种标准品和质控品,用分析缓冲液作为(本底)。在背景孔和所有标本孔中加入25ul分析缓冲液。在本底、标准品和质控品中加入25ul基体溶液。加入25ul标本到适当孔中。涡旋混合瓶,每个孔中加入25ul混合的珠子,在加珠子过程中间歇的摇动装珠子的瓶子,避免沉淀。用封板膜封板,在板的外面上铝箔,放在振荡器上室温(20-25摄氏度)振荡孵育2h或者4℃孵育过夜。轻轻地去掉板里的内容物,洗板两次。在每个孔中加入25ul检测抗体。封板,包上铝箔,在振荡器上室温振荡孵育1h。在每个含有25ul检测抗体的孔中加入25ul链霉亲和素-藻红蛋白。封板,包上铝箔,在振荡器上室温振荡孵育30min。轻轻地去掉板里的内容物,洗板两次。在所有孔中加入150ul鞘液。在振荡器上重悬珠子15min。将板放在flexmap上用xponent运行。保存并使用样条曲线拟合法分析中间荧光强度数据,计算标本中细胞因子的浓度。8细胞计数使用ipp图像分析软件对采集到的免疫组化图像进行阳性细胞计数。不同时间点的标本在400x显微镜下阳性细胞数进行统计,每个时间点取3个样品,每个样品取5个视野,然后取其平均值。使用graphpadprism5软件进行绘图和数据分析。9数据分析及统计学分析所有实验数据均代表3次以上实验结果,以平均值±标准差进行统计。数据均由专业统计软件spss或graphpadprism5进行分析(13.0版),两组数据间采用单因素方差分析处理。p值小于0.05表示具有显著性差异。*p<0.05,**p<0.01,***p<0.001。(三)结果1恒河猴外周血细胞因子、趋化因子和生长因子的动态变化我们利用luminex技术检测了恒河猴外周血清中细胞因子、趋化因子和生长因子共29个因子的动态表达变化。如图9所示,两组动物外周血清炎症因子tnf-α、il-1β、il-1ra、il-15、ifn-γ,趋化因子mcp-1、mip1α、mip1β和生长因子hgf、vegf、fgf-basic于amanitin毒素注射后3天内均略微升高,之后在msc-组随肝功能恶化而持续上升,第4-5天时显著高于msc+组。在msc+组,上述因子表达水平在各时间点均无显著变化或仅有轻微升高,第5天时迅速回落,至第7天已恢复至正常水平。msc-组il-6在第1天时迅速达到峰值,较毒素诱导前升高达100余倍,也高于msc+组34倍。之后il-6水平急转直下,但至第5天时仍6倍高于msc+组。两组动物il-2、il-8、il-10、il-12、g-csf、egf、rantes、eotaxin、mif、i-tac、mdc、mig、ip-10等因子在各时间点两组的表达水平无显著性差异。il-4、il-5、il-17和gm-csf等因子低于检测阈值,未能测出结果。2单核细胞活化检测外周循环中炎性细胞因子检测结果提示,毒素输入引起了恒河猴全身性的炎症反应。外周血il-6在第1天就急剧升高,提示其在alf启动及进展中可能起重要作用。多种细胞均可分泌il-6,例如活化的t细胞、单核细胞甚至成纤维细胞。然而,急性肝损伤发生后,首先激活肝内巨噬细胞即库否细胞(kcs),后者成为分泌il-6的主要细胞群。kcs的活化及分泌il-6是促进肝脏修复的重要保护性因素。但kcs的过度活化也成为促进局部和全身系统性炎症反应的关键环节。il-6在alf早期的急剧升高提示kcs的大量动员活化,这可能是毒素导致alf病情进展的关键环节。cd163是血红蛋白清道夫受体,特异性表达于活化的巨噬细胞膜。血清中的可溶性cd163(solublecd163,scd163)的水平能反映巨噬细胞活化状况,也可间接反映肝衰竭病情严重程度[60-62]。我们用elisa分析了两组动物血清中scd163水平。在第1天时msc-组scd163就急剧升高,而msc+组仅略微升高,两组相比具有极显著性差异(p<0.001),这也提示了msc-组巨噬细胞的显著活化(图10)。肝脏内的kcs是体内最大的巨噬细胞群,早期急剧升高的il-6和scd163提示kcs的大量活化。cd68是成熟kcs的特异标志物。免疫组化染色结果显示在msc-组,直到第3天才见到明显增多的cd68阳性细胞,尽管其kcs胞浆丰富,细胞体积较大,但与msc+组相比,其数量并无显著性差异(图11)。肝脏巨噬细胞的增多,一部分可能为kcs分裂增殖,另一部分可能来源于循环中的单核细胞(circulatingmonocytes,c-mos),受到ccr2(单核细胞趋化蛋白mcp-1受体)等信号趋化而迁移到肝损伤局部,分化为kcs。我们检测了肝脏内kcs增殖情况,如图12所示,肝脏内罕见cd68+ki67+细胞,表明kcs的增殖并不活跃。接下来我们利用免疫组化检测了由循环中迁移入肝脏的单核细胞(c-mos)。mac387是外周血迁移入肝而又尚未分化成熟的单核-巨噬细胞。结果显示早在第2天,msc-组即可观察到大量mac387+细胞聚集于肝脏坏死灶周围,第3天继续增多,至第4天时随着肝实质的大量丢失其形态发生改变(图11a,b);而msc+组在各时间点均未出现mac387的显著增多(图11a,c)。以上结果提示,肝内巨噬细胞的大量活化,绝大部分是从c-mos募集而来。单核细胞趋化蛋白1(mcp-1)与特异性趋化因子受体2(ccr2)结合后,激活单核-巨噬细胞,被认为具有促进炎症反应的作用,高表达ccr2的单核细胞被视作活化的单核细胞。我们采用流式细胞术检测了外周血cd14+cd16+ccr2+的单核细胞。毒素诱导后第1天,尽管我们前面未发现单核细胞总数升高,但msc-组动物外周血cd14+cd16+ccr2+阳性的c-mos比例显著升高,且远高于msc+组(图13)。为明确活化的c-mos是否是il-6的来源,我们采用rt-pcr检测了其il-6mrna的水平。结果显示,从msc-组分选所得的c-mos其il-6的mrna显著性升高(图14)。cd45是炎症细胞的共同表面标志,我们进而通过cd45免疫组化检测了两组动物第3天时肝内炎性细胞浸润情况。与msc-组相比,msc+组局部炎性细胞的浸润被明显抑制(图15)。本部分结果表明,msc-组肝内巨噬细胞主要是由ccr2趋化的c-mos迁移至肝脏聚集而成,而il-6在第1天时急剧升高,正是来自于cd14+cd16+ccr2+的c-mos。huc-mscs输入后,显著减少了活化的c-mos并抑制其分泌il-6,进而有效地阻断了系统性炎症反应的过度放大,逆转了毒素引起的恒河猴alf的发生。3预防性输入huc-mscs有效抑制毒素诱导的肝损伤huc-mscs的输入强有力地抑制了c-mos活化进而改善了毒素引起的alf。我们进而探讨了预防性给与huc-mscs是否同样能保护肝细胞免于毒素损伤。我们在毒素诱导建模前的-9,-8和-7天分别通过外周静脉给予1×107个huc-mscs,共计3×107huc-mscs,作为预防组(pre-msc+)。实验流程如图16所示。结果显示,预防组动物除了在毒素给予后的前两天食欲有所减退,之后始终保持良好的生理和精神状态,动物无一死亡。肝功生化指标显示,与毒素诱导前相比,pre-msc+组第2天时酶学指标、胆红素和血氨水平几乎没有发生改变,第5天时alt仅为501u/l,胆红素和血氨水平与正常水平相当(图17a)。其肝功能指数明显好于msc-组,甚至好于msc+组。第3天时肝活检组织的he染色结果显示为正常的肝小叶结构,cd68和mac387阳性细胞极少(图17b)。外周血清scd163水平检测也显示第1天时pre-msc+组仅较注射毒素前略有上升,但无统计学差异,表明scd163也被显著抑制(图18)。同样,流式细胞术检测pre-msc+组cd14+cd16+ccr2+单核细胞比例,也显示其外周血活化的c-mos接近于正常健康动物,并且这群单核细胞的il-6mrna表达水平也不升高(图19)。同样,提前输入huc-mscs并未引起外周血白细胞数目和treg等淋巴细胞亚群比例发生变化(图20)。我们又采用luminex液相芯片检测了细胞因子水平。与健康对照比较,从第2天到第5天,pre-msc+组动物外周血清中炎症介质,如il-1α、tnf-α、il-1β、ifn-γ,趋化因子,如mcp-1、mip1α、mip1β等几乎无显著改变或仅有轻微升高,外周血il-6亦被显著抑制。某些炎性因子如il-1α等较huc-msc+组更低(图21)。该结果提示预防性给予huc-mscs似乎能更显著地抑制局部及后续的系统性炎症反应。(四)讨论通过流式我们分析了外周血cd14+cd16+ccr2+单核细胞亚群,结果证实alf猴c-mos被显著活化,高表达il-6mrna,成为血清中il-6的主要来源。huc-mscs能明显抑制c-mos的活化和迁移过程,并抑制其产生il-6。同时,我们的研究也证实huc-mscs治疗和预防alf的的靶细胞正是c-mos。(五)结论外周血输入的huc-mscs通过抑制c-mos的过度活化及产生il-6,从而显著抑制继发性系统性炎症反应,使恒河猴免于致死性alf。实验结果说明,通过抑制外周血单核细胞的活化,可以有效治疗和预防急性肝损伤,其中,huc-mscs即可以有效抑制单核细胞的活化,具体地,可以抑制活化标志物ccr2的表达,同时抑制il-6的分泌,说明huc-mscs是一种外周血单核细胞的活化抑制剂,可以通过抑制外周血单核细胞的活化,进而治疗和预防急性肝损伤。综上,本发明实验从基因水平证实了抑制单核细胞的活化能够有效治疗和预防急性肝损伤,再以脐带间充质干细胞验证了抑制单核细胞的活化的确能够有效治疗和预防急性肝损伤,说明单核细胞活化抑制剂可以有效治疗和预防急性肝损伤,可以作为治疗和预防急性肝损伤的药物使用,其中,单核细胞活化抑制剂可以是il-6基因敲除型单核细胞,也可以是脐带间充质干细胞,还可以是其他抑制单核细胞活化的物质,如il-6基因沉默剂。本发明为临床治疗急性肝损伤提供了一种新的选择,应用前景优良。sequencelisting<110>四川大学华西医院<120>外周血单核细胞活化抑制剂在制备治疗急性肝衰竭的药物中的用途<130>gy026-17p1109<160>7<170>patentinversion3.5<210>1<211>20<212>dna<213>il-6pcr引物f<400>1tcgagtatctgacaatgtgg20<210>2<211>21<212>dna<213>il-6pcr引物r1<400>2tagccagaagttccaaattgg21<210>3<211>22<212>dna<213>il-6pcr引物r2<400>3atagcctgaaaccaactgtgcc22<210>4<211>20<212>dna<213>il-6qrt-pcr引物f<400>4atgaggacacttgcctggtg20<210>5<211>20<212>dna<213>il-6qrt-pcr引物r<400>5gctggcatttgtggttggtt20<210>6<211>21<212>dna<213>gapdhqrt-pcr引物f<400>6tcgagagtcagccgcattttc21<210>7<211>20<212>dna<213>gapdhqrt-pcr引物r<400>7ggaacttgccatgggtggaa20当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 干扰素调节因子5(IRF5)及其抑制剂在治疗心肌肥厚中的应用的利记博彩app与工艺
- 作为因子XIA抑制剂的环状P1连接体的利记博彩app与工艺
- 能够结合组织因子途径抑制剂(1‑161)上的两个表位的抗体的利记博彩app与工艺
- 针对组织因子途径抑制剂(TFPI)的优化的单克隆抗体的利记博彩app与工艺
- 一种作为自分泌运动因子抑制剂的咔唑类化合物及其制备方法和应用与流程
- 外周血单核细胞活化抑制剂在制备治疗急性肝衰竭的药物中的用途的制造方法与工艺
- 对NEDD8活化酶(NAE)抑制剂的反应的生物标记的制造方法与工艺
- 一种肺癌调节因子SRSF5及其抑制剂与应用的制造方法与工艺
- 自分泌运动因子抑制剂的制造方法与工艺
- 一种BCL‑2抑制剂Venetoclax的合成方法与流程